
 welcome�͔쉮com
��
welcome�͔쉮com
�� 
�ŏI�X�V��2005/05/27
|
|
|
|
|
�ŏI�X�V��2005/05/27 |
|
�^���p�N��(�`����)�@�^���p�N���@Protein �����̂��\�������v�ȍ����q�����B�A�~�m�_�������Ȃ��������́B1838�N�ɔ������ꂽ�B�זE�̎听���ł���A�������ۂɐ[���������������Ă���B�v���e�C��protein�Ƃ�����́A�M���V�A��́u���̂��́v�Ƃ����Ӗ���proteios�ɗR������B �^���p�N���̕��q�ɂ́A�����g�D��т����鐅�ɂƂ��Ȃ��@�ۏ�A�זE���ɓ��ߐ��������������Ӕ������Ђ����������ɂƂ��鋅��܂ŁA���܂��܂Ȑ����̂��̂�����B���q�ʂ͐��炩��S���ȏ�̋��啪�q(�����q)�ŁA�킲�Ƃɓ��قŁA��̊튯�ɂ�鍷������B �l�Ԃɂ́A3����̃^���p�N����������̂Ƃ݂��邪�A���e�����イ�Ԃ�ɂ킩���Ă�����͖̂�2%�ɂƂǂ܂�B�H�����̃^���p�N���̓A�~�m�_�ɕ�������ċz�����ꂽ�̂��A�זE�̍\�z�ޗ��ƂȂ邪�A���܂������͕̂����ɂ���ăG�l���M�[���������A�Y��������1g������4cal�Ƃقړ����M�ʂ����݂���(�� ���)�B �����ƍזE�̈ێ��@�\�̂ق��ɁA�^���p�N���͋ؓ��̎��k�ɂ��֗^���Ă���B�C���X�������̑��������z���������A�܂��A���ׂĂ̏����_�f���^���p�N���ł���B�Ɖu�n�̍R�̂��^���p�N���ł���B�w���O���r���́A�����ێ��ɕK�v�Ȏ_�f��S�g�ɂ͂��ԃ^���p�N���ł���B�^���p�N���́A���ׂ���`�q�ɂ���Ă��̃A�~�m�_�̕��я�(1���\��)�����߂��Ă���B �h�{
�����ێ��ɕs���ȃ^���p�N������������ɂ́A 20��ނ̃A�~�m�_�����ꂼ����̊����ŕK�v�ł���B�A�����������ɂ���āA�K�v�ȃA�~�m�_�����ׂāA���f�A��_���Y�f�Ȃǂ̕��i������肾��netodojou.htm�邪�A�����ȂǑ����̐����́A�ꕔ�������������邱�Ƃ��ł��Ȃ��B����ȊO�̃A�~�m�_(�K�{�A�~�m�_)�́A�H������ێ悷��K�v������B�l�Ԃ̏ꍇ�A���N���ێ�����ɂ́A 8��ނ̕K�{�A�~�m�_���K�v�Ƃ����B���C�V���A�C�\���C�V���A���W���A���`�I�j���A�t�F�j���A���j���A�g���I�j���A�o�����ł���B�����͂��ׂāA�A���̎�q�Ȃǂ��^���p�N���ɂӂ��܂�Ă��邪�A�l�ԂɂƂ��Ă̓��W���ƃg���v�g�t�@�����s���������Ȃ̂ŁA�h�{�w�̐��Ƃ́A�H���ɓ��E���E�����Ȃǂ̕K�{�A�~�m�_�����イ�Ԃ�ɂӂ��ޓ������^���p�N���̕⋋�������߂Ă���B�������^���p�N�������イ�Ԃ�ɐێ悵�Ă��鍑�ł́A�K�{�A�~�m�_�̕s���͂قƂ�ǂ݂��Ȃ��B�M�уA�t���J�̎q���ɂ݂���N���V�I���R���Ƃ����a�C�́A�K�{�A�~�m�_�̌��R�ɂ��ƂÂ����̂ł���B���l�̏ꍇ�A�^���p�N���̕K�v�ʂ͑̏d1kg������1��0.79g�Ƃ݂����Ă���B������c���ł́A�������}���Ȃ��߁A���̐��l�����ꂼ��2�{�A3�{�Ƃ���(�� �h�{)�B�^���p�N���̍\�� �^���p�N���̂����Ƃ���{�I�ȍ\���ł���1���\���́A�A�~�m�_�̒����z��������B���ɂ�������X�̃A�~�m�_�̔z��́A�^���p�N�����q�̍\���ɂ��܂��܂ȉe���������炷�B���f�����A�W�X���t�B�h����(�������q���ӂ��ރV�X�e�C����2���q�A�Ȃ��肠��)�A�����d�ׂ̈��������A�a�����Ɛe�����̌����Ȃǂɂ���āA�^���p�N�����q�͉Q����ɂ܂�����A���肽���܂ꂽ�肵�āA�A���t�@�点��Ƃ��x�[�^���Ƃ�����2���\��������B ���q�S�̂��A����^���p�N���Ȃǂ̂܂Ƃ܂����������Ԃ��A3���\���Ƃ����B�^���p�N�����A�w���O���r���⑽���̍y�f�ł݂���悤�ɁA2��葽���̃|���y�v�`�h�������܂��ă^���p�N�����q�Ƃ��Ċ�������ꍇ�ɂ́A4���\�������Ƃ�����B���̂Ƃ��A�X�̃|���y�v�`�h�����T�u���j�b�g�Ƃ����B �@ �@
�^���p�N��(�`����)�@����ς����@protein �^���p�N���́C�����̂��\����������Ƃ���{�I�ȕ����ł���C���܂��܂ȏd�v�ȓ��������Ă���B�זE���O�Ɍ������X�̍\���́C��Ƃ��ă^���p�N���ɂ��`������邵�C�����ɕK�{�̉��w������G�}����y�f�́C�^���p�N���łł��Ă���B�����āC���̉^���C�_�o�n�̊����C�����̗A���C�Ɖu�����Ȃǂ��^���p�N�����s���Ă���B���Ȃ킿�C�^���p�N���̑� 1 �̓����Ƃ��āC���̋@�\���������邱�Ƃ��ł���B �@�܂��C�^���p�N���͂��̂��̂̐�����ɌŗL�̂��̂ł���C������̓����̓^���p�N���ɂ�茈�܂��Ă���B�^���p�N���́C�l�Ԃ��͂��߂Ƃ��Ă��ׂĂ̓����̉h�{���Ƃ��ďd�v�Ȃ��̂ł���
(�^���p�N���̉h�{�I���ʂɂ��Ă� �@DNA������ 20 ���̓�d�点��̎��Ƃ����ψ�ȍ\�������̂ɑ��C�^���p�N���̗��̍\���͎�ނɂ��C���̐܂肽���܂�����S�̂̊O�`���Ђ��傤�ɑ��ʂł���B DNA �͂ǂ���Ƃ��Ă݂Ă����̒����E�����E�����Ƃ�������ꂽ����̋@�\���������Ȃ��̂ɑ��C�^���p�N���͋@�\�I�ɕ������Ă���C��ނɂ��قȂ邳�܂��܂ȋ@�\���ʂ����B���ꂪ�\����̑��l���ɂ��\��Ă���̂ł��낤�B DNA �ɂ��Ă͈�ʘ_�ő����̂��Ƃ�����ł��邪�C�^���p�N���͊e�_�����Ȃ�������C���̖L�����Ɛ�����������ł��Ȃ��B���̂悤�ɁC�^���p�N���̑� 3 �̓����́C���̑��l���ł���B �@�זE�́g�����Ă���h�����ł������`���́C�����������Ǝ�v�ȕ����̓^���p�N���łł��Ă���B���������āC100
�N�ȏ�O�̐l�X���C�����̐_��������ړI�ŁC�܂��^���p�N���̌����ɒ��肵���͓̂��R�ł������B�^���p�N�����Ӗ�����
protein �Ƃ����ꂪ�M���V�A��Łg�� 1 �́h�Ƃ����Ӗ�������
pr�S��teios
�ɗR������̂��C���̂悤�Ȕw�i����l����Ɨ����ł���B�������C19
���I���� 20
���I�����̊w�E�ł́C�܂����낢��ƍ��������l�����݂�ꂽ�B���Ƃ��C �@ �\ 1 �Ɏ����^���p�N�������̗��j�́C����C�����̌�����l���������X�ɒ�������Ă䂭���j�ł�����B�����ē��B�������݂̃^���p�N���ς͎��̂Ƃ���ł���B�^���p�N���ɂ͂Ђ��傤�ɑ����̎�ނ�����C�킲�Ƃɑ傫���E�`�E���w�g�����قȂ�B�������C���̃^���p�N�������グ�Ă݂�ƁC����͈��̑傫���E���̌`�E���̉��w�\���������Ă���B���̍\���́C���w�\�� (�A�~�m�_�z��)�E���̍\���Ƃ��Ђ��傤�ɕ��G�ł���C�P���ȍ\���̌J�Ԃ��ɊҌ����邱�Ƃ͂ł��Ȃ��B���G�ȉ��w�\�����܂��������ɍ����ł���̂́C��`���ɏ]���č���邩��ł���B�܂��C���G�ȗ��̍\���́C�^����ꂽ���w�\���͈͓̔��Ŏ��R�G�l���M�[���ŏ��ɂȂ�悤�Ɍ��܂�B �\�\����ł́C���̃^���p�N���ς͍ŏI�I�Ȃ��̂Ȃ̂��낤���B�܂��C�����ɂ͂ǂ�Ȗ�肪�c����Ă���̂��낤���B�܂��C���݂̒m�������Ă���C�Ō�ɂ܂����̖��ɖ߂��Ă݂悤�B �@�^���p�N���̍\���͈ꎟ����l���܂ł̒i�K�ɕ����ċL�q����邱�Ƃ������B�ꎟ�\���Ƃ͂����鉻�w�\���ŁC�A�~�m�_���ǂ̏����Ɍ������Ă��邩�������B��ʂɃ^���p�N���̓v�������c�� ( �} 1 ) ���O�Ƃ��āC���̂悤�ȍ\�����ŕ\�����B
�����ŁC���̎��̍��[�̓A�~�m���[�C�E�[�̓J���{�L�V�����[�C�|
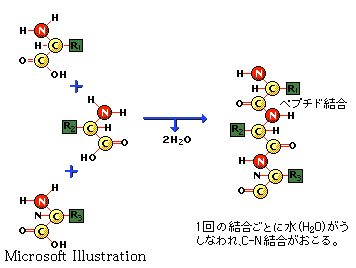
CO �| NH �|�� �@�\���Ƃ͑����̃^���p�N���Ɍ���������镔���I�������I�ȗ��̍\���̂��Ƃł���B�^���p�N�����̌��q�Ԍ����̒����ƌ����p
(��̌��q����o�錋���Ԃ̊p�x)
�́C�ꎟ�\���ɂ�炸�قڈ��Ƃ݂Ȃ��Ă悢 ( �}
2-a )�B���������āC���̍\���ő傫���ω�����̂́C�����̂܂��̓�����]�p�݂̂ł���B�卽��
NH �| C������ C�����|
CO �̂܂��̓�����]�p�����ꂼ��ӂƃՂŕ\�� ( �}
2-b )�B�卽�̂�����̌����ł��� CO �| NH
�̂܂��̓�����]�p�́C�ق� 180 �x (�g�����X)
�ɌŒ肳��Ă���Ƃ݂Ȃ��Ă悢�B (�ӁC��)
�̓�����]�p�̑g�ɂ́C�������̃G�l���M�[�̒Ⴂ����������
( �}
2-c )�C���ꂪ�A�������A�~�m�_�c��Ɏ��������Ƃ��C�\���Ƃ��ĔF�������B
�@�|���y�v�`�h�� 1 �{�̑S���̍\�����^���p�N���̎O���\���Ƃ����B���S�ȎO���\���́C�卽�Ƒ������܂ޑS���q�̎O�����̈ʒu���W�ŕ\�����B�������C���f���q�̈ʒu�́C�^���p�N���̎O���\�������߂� X ����͖@�ł͕��ʌ����Ȃ��B���ۂɐ}������ꍇ�C�S���q�̈ʒu��`���ƕ��G�����Č��ɂ����Ȃ�̂ŁC �����Y�f�̈ʒu�݂̂��������� ( �} 4-b-I )�C�\����͎��I�ɕ\�������� ( �} 4-b-II ) ����ꍇ�������B�|���y�v�`�h���́C�@�ۏ�^���p�N�� ( �} 5 ) �ł� 1000 ���ȏ�̒����ɐL�тĂ��������邪�C����^���p�N���ł͊ۂ܂��Ē��a 25 �` 60 ���̗��ɂȂ��Ă���B�O���\���̈ꕔ�ɂ͓\���̑g�������悭����C����\���Ƃ������Ƃ����� ( �} 4-a )�B�܂��C�O���\���̒��ŗ��̓I�ɂ݂Ă܂Ƃ܂��Ă���悤�ȕ������h���C���Ƃ������Ƃ����� ( �} 4-b-II �� �} 8-b �ɂ��̗Ⴊ����)�B �@�^���p�N���̂�����͕̂����̃|���y�v�`�h�����琬��B���̏ꍇ�C�|���y�v�`�h���̐��Ƃ����̊Ԃ̊w�I�ʒu�W�����킹�Ďl���\���Ƃ��� ( �} 4-c )�B�O�ʑ̂̃^���p�N�����������邪�C�|���y�v�`�h���̐��� 2�C 4�C6 �ȂNj��������ʂł���B �@�^���p�N���ɔM�∳�͂���������C�n�t�� �@����ł́C�A�~�m�_�z�^����ꂽ�Ƃ��C���̃^���p�N���̎�X�̐������ǂ̒��x�\�����邱�Ƃ��ł��邾�낤���B�A�~�m�_�c��ɂ��C��X�̓\�������₷���C���邢�͍��ɂ������̂����邱�Ƃ��o���I�ɒm���Ă���
( �\
2 )�B���������ƂɁC�ꎟ�\������\����\�����邱�Ƃ͂��Ȃ萳�m�ɂł��邪�C�O���\���̗\���͂܂�����B�O���\���ł͐e����
(�ɐ�) �̃A�~�m�_���\�ʂɁC�a���� (��ɐ�)
�̃A�~�m�_�����q�����ɂ���X���������B���q�����ł͌��q�̓A�~�m�_�̌��������Ɠ����x�ɖ��ɋl�܂��Ă���
( �}
6-a )�B�P�ʎ��ʂ̃^���p�N�����Ђ��傤�ɑ��ʂ̗n�}�ɗn�������Ƃ��̑̐ϑ�����Δ�e�Ƃ����B�Δ�e�͖��x�̋t���ɋ߂����C�^���p�N���ł͂ӂ�
0.69 �` 0.75 �@��������V�R�̏�Ԃł́C�^���p�N���̗��̍\���͂��܂��܂ȔL�����ɂ����艻����C�G���^���s�[H���������Ȃ��Ă���B����ɑ��C�ϐ���Ԃł͂����̌����͂قƂ�ǐ�Ă��邪�C���̑��ɑ����̎��R�x���l�����ăG���g���s�[S�̑傫�ȏ�ԂɂȂ��Ă���B����M�͊w�ɂ��ƁC�M�u�X�̎��R�G�l���M�[G�̏�������ԂقLj���ł���C
G��H�|TS�Ƃ����W���������邱�Ƃ��m���Ă���B���������āC��Ή��xT�̒Ⴂ�Ƃ��͓V�R�̏�Ԃ��C�܂������Ƃ��͕ϐ�������Ԃ�����ɂȂ�B�܂��C�M�ϐ��̂Ƃ����鉷�x�����ɂ��̓�̏�Ԃ͋}���ɓ]�ڂ���B�V�R�̍\�������艻����L�����Ƃ��āC
���f�����C�Ód����
(����)�C�t�@���E�f���E���[���X��
(���q�ԗ�)�C
�@�^���p�N���̐������́C��`���ɏ]���ăA�~�m�_�������悭�Ȃ����Ƃł���B���̋@�\�͂��Ȃ蕡�G�ŁC�����̐�����K�v�Ƃ���B
DNA ���� 1 �{�𒒌^�ɂ��Ďʂ��Ƃ�ꂽ���b�Z���W���[
RNA (mRNA �Ɨ��L)
�̉���͎O���������ēǂ܂�C���̎O�g (�g���v���b�g�܂��̓R�h���ƌĂ��)
�ɑ��ăA�~�m�_����Ή�����B���̑Ή��̂���������`�Í��\�Ƃ���
( �@�^���p�N���͍�����ɏC�������ꍇ�����Ȃ葽���B���̈��̓y�v�`�h�����̐ؒf�ł���B�g���v�V���Ȃǂ̃^���p�N�������y�f��C���V�������Ȃǂ̃y�v�`�h�z�������ɂ́C�ؒf���ď��߂Ċ����̏o����̂������B�����̖��^���p�N���͍������C�A�~�m���[�ɃV�O�i���y�v�`�h�ƌĂ�镔�������B���̕����̓^���p�N���������ɐ������g�ݍ��܂��Ƃ��ɕK�v�����C��ɐ�����B�܂��ꕔ�̓����E�C���X�ł́C�Ђ��傤�ɒ����|���y�v�`�h�������ɐؒf����C���̂��̂��ʂ̊�������^���p�N���ɂȂ�B �@������A�~�m���[���C�����邱�Ƃ�����B�����ł̓��`�����C���_���C�����_���ȂǁC�A�~�m���[�ł̓A�Z�`�����C�s���O���^�~�����Ȃǂ��N����B�����_���ɂ�芈���̒��߂���y�f�͑����B�܂��C�����̔�����`�q�ɂ́C�^���p�N���������_�����銈��������C���ڂ���Ă���B�L���Ӗ��ł́C���^���p�N���ɂ����铜�̌�����C�y�f�^���p�N���ւ̕⌇���q��
(�`�g�N����c�̃w���Ȃ�)
�̌������C���̕��ނɓ��邾�낤�B�܂��C��̃V�X�e�C���c���
SH ��_������C���q���� �@�Ђ��傤�ɑ����̎�ނ��琬��^���p�N���ނ��悤�Ƃ��鎎�݂͂��낢�날��B�|���y�v�`�h���݂̂��琬����̂� �@�^���p�N���@�\�̍�p�@�\�Ɋւ��ẮC�ǂ̂悤�ȓ������������邾�낤���B���̑�
1
�͌����ɂ����闧�̑���ł���B�^���p�N���ƃ^���p�N���C����у^���p�N���Ƒ��̕��q�Ƃ̌����ʂł́C�ʉ����݂��ɗ��Ԃ��ɂȂ��Ă���C�]���Ȃ����܂��ł��Ȃ��悤�ɂȂ��Ă���B����͌����̓��ِ������߂�������ʂ����Ă���B
�}
8-c �̍R�� (�Ɖu�O���u����)
�ƍR���̌����͂��̈��ł��� (�Ȃ� �}
8-a �ɖƉu�O���u�������q�̃|���y�v�`�h��������)�B�܂��Ƃ��� �@�� 2 �ɔ�����̍œK�z�u������B�y�f��������������Ƃ��������ƁC��̍U�����ׂ������̋ߖT�ɍy�f�̔����� (Ser�C His�CCys�CAsp�CGlu �Ȃǂ̑����C�����C�I���C��y�f�Ȃ�) �����݂��āC�����̌��������߂Ă���B���q�Ԃ̌����ʂł��P�ɗ��̓I�ȓʉ������łȂ��C������ׂ����ɂ�����ׂ�������卽���z�u����C���ƕ��̓d�ׂɂ�錋�� (����) ��| NH (�܂��́| OH) ������C �� O �ɂ�鐅�f�����Ȃǂ��`���ł���悤�ɂȂ��Ă���B���̎���� �} 6�]c�Cd �Ɏ�����Ă���B �@�� 3 �ɂ͌����̗U���K�� induced fit ����������B�n�t���ł̃^���p�N���̗��̍\���́C�킸���ɈقȂ鑽���̍\���̊Ԃ��ڂ�ς���Ă�炢�ł���B�Ƃ��낪����Ɋ�Ȃǂ���������ƁC�������\�ȍ\���݂̂ɌŒ肳���B���̂��߁C���ʓI�ɂ͊���y�f�̍\���������ɓK�����悤�ɕς������ƂɂȂ�B �@�����đ� 4 �� �@�ȏ�͂�����x�����ɂ��m�F���ꂽ�@�\�����C�����Ƃ��Ă͑��ɂ��������̋@�\���l�����Ă���B���Ƃ��C�^���p�N���ɂ͑����̉𗣊��d�C�o�Ɏq��������邪�C�����̕��z�ɂ�芈�����S�ɋ����d�ꂪ�����C���ꂪ��ɂ����čy�f�����������Ă���Ƃ�����������B�܂��C�n�t���Ń^���p�N�����q�͎�X�̐U�������Ă��邪�C���̂����̈�̐U�����g���Ċ��H���������Ă���Ƃ�����������B����ɁC�n�}���q���y�f�ɏՓ˂���ƁC���̃G�l���M�[�͂��̐U���ɏW���I�ɏW�߂���Ƃ����l��������B���ɂ́C�y�f�͊��ǂ������ĕ߂炦��̂��Ƃ����l������B�^���p�N���͂Ђ��傤�Ɍ����悭�d�������邾���łȂ��C���̕ω��ɕq���Ɏ������C�܂��ɐ����Ă���悤�ł���B���ꂪ�����҂̑z���͂��h�����C��X�̍l���ށB���̈Ӗ��ł̓^���p�N���ς͌���ł��܂��h�ꓮ���Ă���B�����̍l���̒��ɂ́C�����C�ے肳�����̂�����ɈႢ�Ȃ��B�������C�����ł͂����Ɛ��I�ȍ�p�@�\�����������\�����\���ɂ���Ƃ����悤�B �@�^���p�N���̌����@�� �\
4 �ɂ܂Ƃ߂Ă���B�^���p�N���̎�����F�����Ƃ��ẮC
�r�E���b�g�����C�L�T���g�v���e�C�������C�j���q�h���������C�~���������Ȃǂ�����B�܂��^���p�N���̒�ʂ́C�r�E���b�g�@�C���[���[�@�Ȃǂ̔�F��ʖ@�̂ق��C���O�z���@�C�����d�ʑ���C���f�̒�ʁC�����������ăA�~�m�_�̒�ʂȂǂɂ��s���B�ꎟ�\���́C���w�I�E�y�f�I�ɒf�Љ����Ă���f�Ђ̃A�~�m�_�z��� �@���낢��Ȑ������瓯���^���p�N�������o���āC���̃A�~�m�_�z����ׂ�Ɛi���̓��������킩��B�Ȃ��Ȃ�C�A�~�m�_�z��̈Ⴂ�́C�킪������Ĉȗ��̓ˑR�ψق̒~�ςɂ��̂ŁC�i���̎��Ԃ��v�鎞�v�ƂȂ邩��ł���B
�}
9-a �� �@�l�H�I�ɃA�~�m�_���Ȃ��ă^���p�N�������w�������邱�Ƃ͂܂�����C���ʂȋZ�p�ƃ^���p�N���̎�ނɍ��������@���K�v�ł���B����ɃA�~�m�_����Ȃ���Ƃ��̐��m����
99 ���Ƃ��Ă��C 100 �Ȃ���� 63 ���C300 �Ȃ����
95
�����܂��������Y���ɂȂ��Ă��܂�����ł���B�������C�A�~�m�_��
10
���x�Ȃ����y�v�`�h�́C�ő������@�Ŕ�r�I�e�Ղɍ����ł���B����͐���n�}������e�Ղɂ��邽�߂ɁC�����̏�ŃA�~�m�_���J���{�L�V�����[�����珇�ɂȂ��ł������@�ł���B����Ƃ��ẮC �@���݁C�\�������ł͈ꎟ�\������̔��ʉ��C���^���p�N����\���^���p�N���̎O���\������Ȃǂ������ȖڕW�Ƃ���Ă���B�^���p�N���̍�p�@�\�Ɋւ��ẮC���̍\�������Ă̐��_����������Ă̓w�͂��C�������ԑ̗̂��̍\������Ȃǂ𒆐S�ɐi�߂���ł��낤�B�^���p�N���̉^���Ɋւ�������Ɨ��_�v�Z���狻�����錋�_�̏o��\��������B�܂��ꎟ�\���̃f�[�^������ȗʒ~�ς����̂ŁC���ꂩ�炢�낢��ƗL�p�ȏ������o�����Ƃ��s������B �@���������̖ڕW�Ƃ��ẮC�]�ދ@�\�����^���p�N����v���邱�Ƃ��l������B����ɂ͈ꎟ�\�������ċ@�\��\�����邱�ƁC����т��̋t���K�v�ɂȂ�B���_�I�ɉ\�ȃ^���p�N���̐��́C����ɃA�~�m�_ 300 �c��Ƃ��Ă� 20300�� 10400�Ƃ������V���w�I�����ɂȂ�B����́C�n���̒a���ȗ��o�������^���p�N����̑����Ɣ�ׂĂ�����Ȃ��傫���B�n��̐����ɂ͂Ȃ��܂������V�����^���p�N���ŁC�L�p�ȋ@�\�������̂��������݂��邱�Ƃ��\�z�����B �@ |
![[���w��]](../../gazou/20902801.GIF)